«Культура чая в СССР». Тбилиси, 1989
ЧАСТЬ 1. МИРОВОЕ ПРОИЗВОДСТВО ЧАЯ И ТРЕБОВАНИЯ РАСТЕНИЙ К ПРИРОДНЫМ УСЛОВИЯМ
Ботанико-морфологическая характеристика чайного растения

Происхождение чая
Вопрос о происхождении чайного растения долгое время считался не изученным и спорным. Новые гипотезы происхождения учеными различных стран выдвигаются и в наше время. Так, специалисты Китая, изучая закономерности распределения растений на земном шаре полагают, что родиной чайного растения является провинция Юньнань (Юй-Де-Цзюнь, 1956). В подтверждение этого приводятся данные китайских ученых Фоен-Шао-цю, Су-Чжен, Ли-Лянь-бяо, показывающих крупные размеры диких чайных растений, произрастающих в провинциях Юньнань и Гуйчжоу. Чжуан Вань-Фан (1959) выдвинул следующую гипотезу распространения родичей чайного растения.
В провинции Юньнань берет начало ряд крупных рек. Здесь начинаются протекающая затем по Вьетнаму река Юаньцзян, текущая в Лаос и Камбоджу река Ланьцанцзян, реки Нуцзян, Саэрвэньцзян и Лунчуаньцзян, текущие по Бирме.
При обследовании бассейнов этих рек повсеместно были обнаружены дикорастущие чайные деревья, очевидно, что они выросли из семян, принесенных в эти места течением рек. Постепенно чайные растения распространились по отрогам гор; большую роль в распространении этих растений в доисторические времена мог сыграть человек. Таким образом, район распространения чая, как пишет автор постепенно все более расширялся.
Дальнейшие исследования ботаников и морфологов ряда стра показали, что дикорастущие чайные растения, кроме Китая, произрастают в северо-восточной Индии (Ассам), в лесах Лаоса, Кампучии, Тайланда, Бирмы, Вьетнама, Бутана. Распространены они в основном в долинах и на водосборных склонах восточной части Тибетского плоскогорья. Некоторые ученые Японии придерживаются мнения, что родиной чая является не только Китай, но и Япония, где на островах Сикоку и Кюсю также встречаются дикие родичи чайного растения. Такого же мнения придерживался и А. Н. Краснов (1897), который после личного знакомства с Японией писал: «Виденные мною местонахождения чая на острове Сикоку, близ города Кочи, не оставляют никакого сомнения, что в диких дебрях дубовых лесов чай рос как подлесок искони веков…».
Однако наиболее правдоподобную концепцию о происхождения чайного растения выдвинул в своей работе «Центры происхождения культурных растений» Н. И. Вавилов (1965), согласно которой чайное растение принадлежит к китайскому очагу, расположенному в предгорьях восточной и отчасти южной и юго-восточной окраины Тибетского нагорья. Площадь эта составляет около 2-х млн. кв. км и включает она, кроме Китая, граничащие с ним горные районы Бирмы, Тайланда, Лаоса, Вьетнама, Кампучии, Индии, Бутана и горных склонов острова Хайнань. Такого же мнения придерживался и другой известный советский ученый-чаевод И. В. Палибин (1920), который писал, что в восточной части Азии, на высотах 1500—2100 м над уровнем моря, в девственных лесах, состоящих из широколиственных пород, произрастают дикие родичи чая. Кроме Китая также они распространены в Лаосе, Бирме и Таиланде. Встречаются там мало изученные расы чайного растения и даже особые виды, близкие к чайному кусту, которые местное население использует для приготовления чая. По данным М. Nature (1958) в Бирме, по течению реки Иравади, найден дикорастущий естественный межвидовой гибрид чая и камелии. Хроматографические его исследования подтвердили существование естественного гибрида ассамского чая с камелией Иравадийской. Характерными признаками этого вида являются прозрачные точки на поверхности листа и его клинообразная форма.
Советский ученый К. М. Джемухадзе (1976) в 50—60-ые годы провел большую работу по обследованию дикорастущих чайных растений и их биохимическому изучению. Им открыты впервые во многих провинциях Вьетнама огромные массивы дикорастущего чая: в провинциях Нгиало, Лангшон, Лаокай, Хананг. Он сообщает, что «лесные массивы чая других районов провинции Нгиало находятся далеко от населенных пунктов, они до сих пор сохранили первобытный вид и их местное происхождение не вызывает сомнения».
Во многих горных районах ДРВ в провинции Лангшон, расположенной в северо-восточной части Вьетнама, которая граничит с провинцией Юньнань (КНР), а с востока с Гуанси в уезде Тьентхонг в горах на высоте 600—800 м над уровнем моря, имеется массив дикорастущего чая на площади 15—20 га, другие участки на высоте 1500—1700 м, где площадь его превышает 150—200 га. Здесь высота дикорастущего чая достигает 12—14 м, толщина ствола — 30—40 см. Возраст этих насаждений составляет 200—300 лет.
Эти факты еще раз свидетельствуют, что родиной чайного растения одновременно являются и тропические зоны Северного Вьетнама.
Зона естественного происхождения чая на юго-востоке Китая и в других странах Тибетского нагорья характеризуется в летний период теплым и влажным климатом, благодаря господствующим муссонам. В зимний период здесь прохладно, и в январе температура может опускаться до —3, —5°С.
Средняя годовая температура находится в пределах 16—18°С, в том числе в январе 9°С и в июле 22°С. В провинции Юньнань, где было особенно много естественных зарослей чая, минимальная температура отмечается в феврале, а максимальная — в августе. Среднегодовая температура находится в пределах 4°—5°С. Годовое количество осадков не превышает 1500—1800 мм. Все эти районы в летний период характеризуются высокой относительной влажностью воздуха.
Несколько иные климатические условия Ассама — родины ассамской группы чая. Дикорастущие чайные растения встречаются здесь на высоте 30—125 м над уровнем моря. Вся зона эта имеет типичный субтропический климат. Наиболее холодным месяцем является январь, однако, температура не бывает ниже 15°, —18°С. Самые теплые месяцы — июль и август, имеющие среднюю месячную температуру 27°, —29°С.
Выпадаемые осадки также являются следствием муссонов, среднегодовое их количество находится в пределах 2700—4000 мм. Почвы в зонах происхождения чайного растения в основном красноземные, являющиеся продуктом выветривания глинистых сланцев, гранитов, гнейсов и других кислых пород. Вследствие воздействия на них различных природных факторов часть из них оподзолены. В отдельных районах встречаются бурые почвы, образующиеся в результате выветривания коричнево-красных песчаников и коричневых сланцев, также характеризующихся кислой реакцией.
Резюмируя все сказанное, можно полагать, что Восточная Азия с ее лесистыми районами Тибетского нагорья и соседние с ним горные области Индии, Лаоса, Бирмы, Вьетнама, Кампучии, покрытые лесами субтропического и тропического климата, откуда берут начало реки, направленные с одной стороны в Индийский океан, а с другой — в Южно-Китайское море, и являются очагами видового разнообразия чайных растений. Сравнивая эти страны, можно видеть близость их основных флористических элементов.
Древнейшим центром сохранившейся до нашего времени третичной флоры является и остров Хайнань, где также произрастают дикорастущие чайные растения.
У ботаников, занимавшихся систематикой чая, долгое время не было единого мнения о классификации чайного растения. Так, считалось, что черный и зеленый чай, получаемый числе переработки, относятся к разным видам. К различным видам чайного растения некоторые ученые относили и различные садовые формы чая (И. В. Палибин, 1927—1928). Многие его формы принимались одна вместо другой. И только в 1922 г. известный английский ученый Кохен-Стюарт предложил вид камелиевых разделить на два географических подвида — китайский чай (Sinensis) и индийский чай (assamica). В каждой из них он в свою очередь выделил разновидности. В частности, в китайском чае японский, китайский и крупнолистный китайский, а в подвиде ассамика разновидности Ассам, Лушай, Наса-Хилл, Макипури, Бурма, Шан, Юньнань и цейлонский гибрид. Все эти подвиды в зависимости от мест произрастания и климатических условий делятся на многочисленные формы. И. Н. Клинген в своей известной книге «Среди патриархов земледелия» пишет по этому вопросу следующее: «…Это несчастное растение в течение более 100 лет не имело постоянного места в систематике».
Морфологические и биологические особенности чайного куста
По современной ботанической классификации чайное растение относится к порядку чаецветных (Theales), семейству чайных (Theaceae), роду чай (Thea), виду чай китайский (Sinensis). Видовое название впервые дано известным ботаником Карлом Линнеем в честь греческой богини Tea в 1753 г.
В семейство чайных входит 23 рода и 380 видов, которые произрастают в тропических и субтропических странах земного шара. Почти все представленное семейство чайных — вечнозеленые деревья, достигающие до 30 м высоты или кустарники, лишь два вида родов этого семейства — Стюартия и Франклиния — являются листопадными растениями, про израстающими в районах с тепло-умеренным климатом. По своим биологическим и морфологическим характеристикам чайный куст очень похож на группу камелиевых растений и поэтому и сейчас в нашей и зарубежной литературе отдельные авторы его относят к роду камелии (Camellia). Однако сходство это только внешнее, в листьях и побегах камелии нет кофеина, танина и других веществ, за что прежде всего ценится чай. Кроме того, у чая листья почти сидячие, у камелии — черешковые.

К. Е. Бахтадзе (1971) китайский вид чая характеризует как небольшое растение, достигающее не более 2—3 м высоты с вертикальной или полураскидистой кроной. Листья у этого вида небольшие (4—10 см) с короткими междоузлиями. Поверхность листа твердая, темно-зеленого цвета, с жесткими плоскими краями и четко видным жилкованием. Почки, а также молодые побеги покрыты нежными серебристыми волосками, которые опадают у взрослых листьев (по-китайски «байхао», отсюда и все название — чай, приготовленный из молодых побегов, называется байховым).
Растения ассамского вида на родине достигают 10—18 м высоты, имеют раскидистую крону и штамб до 1 м в диаметре. Произрастает в тропическом поясе. Листья у этого вида очень нежные, овальные, до 20—25 см в длину и 5—7 см шириной с ясно выраженным кончиком и небольшим основанием, слабо пузырчатые, с волнистыми краями, семена крупные, до 20 мм в диаметре. Уже при —3°, —4°С растения подмерзают.
Из разновидностей индийского чая наиболее распространены Ассам, Манипури, Нага-Хилл, Лушай, Шан, Бурма, Юньнань и др. В ряде чаепроизводящих стран встречается индокитайская разновидность чая, являющаяся гибридом между китайскими и индийскими разновидностями. Это растение в местах произрастания достигает 5—10 м высоты с крупными темно-зелеными листьями (15—17 см). Отличается высокой урожайностью и низкой морозостойкостью.
Китайская разновидность чая, широко распространена в Китае, Японии и некоторых других странах с более суровыми климатическими условиями. Кохен-Стюарт (1916), в зависимости от районов произрастания дикорастущего чая выделил в этом виде следующие разновидности: японскую, собственно китайскую и крупнолистную китайскую.
Растения чая собственно китайской группы имеют темно-зеленые кожистые листья средней величины с 6—7 парами «нервов» (жилок). Сорта этой разновидности отличаются большой жизненностью и морозостойкостью, начинают повреждаться только при —10°, —12°С. Форма крупнолистного китайского чая промышленного значения почти не имеет. Листья у таких растений крупные (до 14 см), темно-зеленые; семян они не дают. К. Е. Бахтадзе (1971), полагает, что это естественно возникшая в природе автотриплоидная форма.
Растения чая японской группы низкорослые, с сильно укороченными междоузлиями, мелкими (длиной до 4—5 см) листьями, собирать которые очень трудно. Пластинка листа толстая, кожистая, жесткая, с редким жилкованием, нервация слабая.
По данным С. Stuarta (1916), японская разновидность чая возникла в восточном Китае, откуда она была завезена в Японию.
Все разновидности чая легко скрещиваются между собой и дают много различных гибридов.
Цветки у чая одиночные, иногда собраны в пучки по 3—4 шт. Сидят в пазухах листьев, пониклые, белые с нежным приятным ароматом и ярко желтыми пыльниками, располагаются на ложной кисти, на которой их находится до 5 шт. Венчик состоит из 5 лепестков, находящихся в круге чашечки, и 6 — в круге венчика. Цветки крупные, достигают в диаметре 40—50 мм. Тычинки располагаются вокруг пестика в несколько рядов, количество их не постоянное, в среднем достигает до 100—200 шт. Завязь верхняя, трех-четырех гнездная, столбик трехраздельный. Чашечка несет в себе 5—7 кожистых чашелистиков. Бутонизация у чайного растения на Черноморском побережье Кавказа, как и в Ленкорани, начинается в конце июля, а цветение — в конце сентября и продолжается в течение всей зимы до первых заморозков, от которых цветки быстро погибают. В тропических странах чайный куст цветет круглый год. Закладка цветочных почек начинается в конце июня — начале июля и продолжается до сентября в пазухах листьев. Период с момента их закладки до сбора семян составляет 16 месяцев. Из общего числа цветочных почек не более 2% образуют завязи и дают семена.
Плод у чая трех-, четырех-, реже пятистворчатая коробочка размером 30—40 мм, в начале зеленого цвета с антоциановым налетом, а при созревании — темно-бурого. При подсыхании створки коробочек растрескиваются и семена выпадают на почву. Они округлой формы, темно-коричневые с матовым налетом, иногда приплюснутые. В ядрах содержится около 30—35% чайного масла, поэтому при неправильном хранении масло в них окисляется и семена быстро теряют всхожесть.
В плодовой коробочке находится от 2 до 5 семян. В зависимости от вида и разновидности растений размер их колеблется от 10 до 15 мм в диаметре. На Черноморском побережье Кавказа полное созревание семян происходит в конце октября — середине ноября. Само семя чая желтовато-белого цвета, состоит из 2-х плосковыпуклых семядолей, горького вкуса.
Чайное растение в зависимости от разновидности отличается вертикальной, полураскидистой, раскидистой и пониклой формой ветвления. Вертикальное ветвление характерно для растений индийско-китайской группы. Основные ветви у них слегка прижаты к стволу, придавая кроне пирамидальную форму. Полураскидистая форма характеризуется расположением ветвей под 45° по отношению к центральному стволу, в этом случае крона растения располагается равномерно как по ширине, так и по длине куста. Свойственна она китайским и индийским видам чая.
При раскидистой форме ветвление отходит почти под прямым углом по отношению к центральному побегу или стволу. Такое ветвление имеют отдельные сорта как китайской, так и ассамской группы растений. Пониклая форма чайного растения встречается сравнительно редко, только у китайской разновидности. При этой форме ветвления куст принимает стелющуюся форму. К. Е. Бахтадзе (1948) установила, что урожайность чайного куста в значительной степени связана с габитусом растения: чем шире крона куста, тем больше его листосборная площадь. Крупнолистные растения, как привило, имеют сильные побеги с большим количеством растущих почек.
Анатомические признаки чайного листа сильно варьируют в зависимости от места произрастания, сорта, разновидности, почвенных условий. Размер устьиц, клеток, толщина тканей являются индикаторами жизненности растения. Например, у китайского чая палисадная ткань двухслойная, у индийского — рыхлая, однослойная, у японского чая — сравнительно морозостойкого — трехслойная. Таким образом, у морозо- и засухо- устойчивых сортов чая эпидермис бывает тонкий, но очень плотный, многослойный, со сравнительно большим количеством мелких устьиц, расположенных на оборотной части листа. Для листьев чая характерно наличие 2-х, 3-х слоев клеток, однако в условиях влажного климата клетки 2-го, 3-го слоев преобразуются в губчатую ткань.

В тканях листа индийского чая встречаются крупные клетки — идиобласты, которые служат растению хранилищем запасов воды. Под влиянием ежегодно изменяющегося комплекса климатических факторов, обусловленных чередующимися сменами времен года, растения чая в процессе филогенеза, наряду с возрастными периодами жизненного цикла, выработали своеобразную ритмику роста и развития в годовом цикле, которая дает им возможность проходить ту или иную фазу при наиболее благоприятных климатических условиях. В течение вегетации у чайного куста отмечается усиленный рост побегов и листьев, происходит закладка и развитие генеративных органов. В период покоя прекращается вегетативный рост, замедляется ассимиляция, дыхание, обмен веществ. В каждой фазе своего развития растение чая требует своих особенных условий, выполнение которых в значительной степени обеспечивает нормальное их прохождение. Академик Б. А. Келлер (1940) по этому вопросу пишет: «Человек, применяя то или иное воздействие на среду и растение, может получать карликовые и гигантские формы растений, однолетники превращать в многолетники и наоборот, изменять аромат, вкус и окраску плодов…». В хозяйствах чай возделывается ради получения возможно большего количества листа, поэтому и формируется он в виде куста или шпалеры, которым придается широкая полусферическая поверхность. Соответственно, у него изменяется и схема побегообразования. Д. Т. Патарава (1948), С. X. Пирцахалайшвили (1947) установили, что вместо 5—6 порядков ветвления образуется их на шпалере до 12—14. При сборе чайного листа значительная часть растущих побегов с куста систематически удаляется. Поэтому ежегодная подрезка усиливает вегетативную деятельность растения. Стремясь восстановить нарушенное равновесие, чайный куст выгоняет все новые и новые побеги, напрягая до максимума свою побегообразовательную способность. Вместо моноподиального ветвления, свойственного чаю, растение переходит к симподиальному. Интенсивность фотосинтеза у подрезанных кустов значительно выше, чем у неподрезанных. Это объясняется стремлением растения выработать возможно большее количество пластических веществ для создания новой кроны взамен удаленной. На неподрезанных растениях вегетация начинается раньше на 7—10 дней вследствие того, что почки прошли дифференциацию еще осенью. Однако в дальнейшем рост побегов замещения и урожайность бывают значительно ниже, чем у подрезанных кустов.
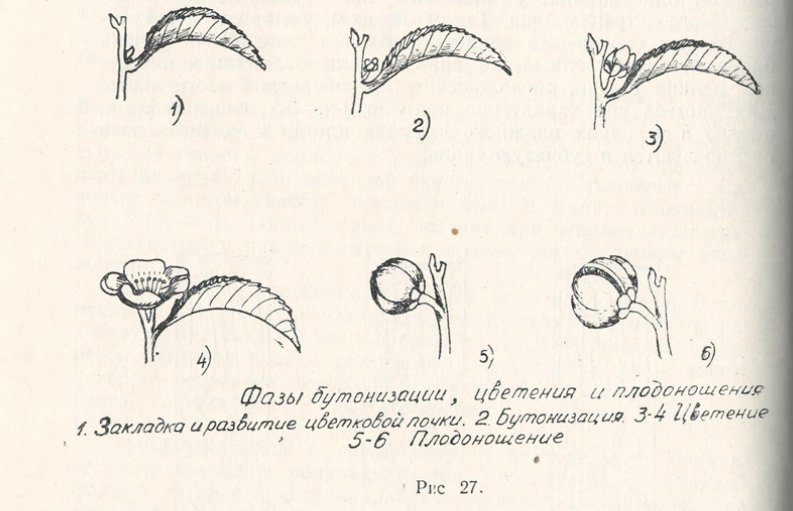
Являясь древнейшим растением земного шара, чай в своем филогенетическом развитии приспосабливался к влажному и жаркому климату тропиков и субтропиков и выработал при соответствующих условиях среды своеобразный ритм роста и покоя.
Вегетация в субтропических районах Грузии и Краснодарского края начинается при среднесуточной температуре воздуха выше 9°—10°С. Первым проявлением которой является набухание ростовых почек и раскрытие почечных чешуек, проходящее в конце февраля — начале марта. В начале апреля уже ясно просматривается развитие побегов 1-го порядка.
Изменение ритма побегообразования зависит прежде всего от условий среды, способов культуры, возраста насаждений. По месту расположения побегов на кусте они подразделяются на верхушечные, пазушные — формирующиеся из спящих почек — и побеги, развивающиеся из придаточных почек. Последние в основном находятся в зоне корневой шейки и начинают расти после тяжелых подрезок или поломов растений. 11о данным К. Е. Бахтадзе интервал между образованием побегов смежных порядков в течение вегетации составляет 12—23 дня. Первый рост побегов в зависимости от погодных условий и сортовых особенностей продолжается с 10—15 апреля по 25—30 июня. Побеги II роста формируются с 25—30 мая, III роста — с 20—25 июня, IV роста — с 20—25 августа и V роста — с 15—20 сентября.
У молодых растений в первые годы их жизни усиленный рост побегов отмечается в продолжении всего вегетационного периода с небольшой ростовой паузой в течение 25—30 дней (в июне месяце). У листосборных плантаций после окончания первой волны роста побегов активно растущая верхушка флешей прекращает рост и глохнет. Происходит период летнего покоя. По окончании этого периода начинается вторая волна роста. Во всех случаях первым после раскрытия почки появляется нежный рыбий лист, а за ним нормальный лист.
Вторая волна роста затухает только с наступлением низких температур — в конце октября или начале ноября. При наличии оптимального ухода за растениями и особенно орошения у молодых растений фаза летнего покоя заметно сокращается.
Спящие почки, образующие новые побеги, находятся в пазухах листьев. Особенно их много у основания побегов. Кроме пазушных и спящих почек на многолетней древесине расположены придаточные почки, которые начинают расти при повреждениях растений или при тяжелой омолаживающей подрезке. Удаление значительной массы листьев при их сборе вызывает нарушение корреляции между надземной и подземной системами, которое и определяет изменение физиологических функций растений — фотосинтез, транспирацию. Стремление растений восстановить срезаемый прирост приводит к сокращению продолжительности его жизни. Поэтому листосборным плантациям особенно важно давать оптимальный комплекс агротехнических мероприятий, который, способствуя увеличению листовой поверхности куста, а соответственно и продолжительности жизни растения чая. Кроме потери листа при сборе урожая часть его подвержена естественному осыпанию. Средняя продолжительность жизни листьев не более 2—3 лет.
Наиболее активными, с хорошо действующим ассимиляционным аппаратом являются молодые однолетние листья, накапливающие к зиме максимальное количество пластических веществ. Более старые листья, наряду со своей ассимиляционной деятельностью, собирают продукты отброса и к концу двух-трехлетнего возраста теряют свою жизнедеятельность и постепенно опадают. В годовом цикле у чая имеется несколько активных периодов сбрасывания листьев, хотя небольшое их опадение происходит в течение круглого года.
Корневая система чайного куста является основной частью растения, которая по своим функциональным задачам, кроме поглощения воды, с растворенными в ней минеральными элементами обеспечивает синтез органических веществ и складирование их в своих запасных вместилищах. У чайного куста на концах корней находятся клетки корневого чехлика, которые предохраняют конус нарастания от механических повреждений. С первых же дней своей жизни сеянец чая образует мощную корневую систему, которая по массе превосходит надземную часть растений. У чая особенно интенсивный рост корней наблюдается на протяжении нескольких лет. К концу первой вегетации у однолетних сеянцев образуется до пяти порядков ветвления с общим количеством 10—12 тысяч корней и суммарной их длиной 75—100 м.
Снаружи корень чайного растения покрыт эпиблемой, за которой расположена первичная кора, состоящая из экзодермы, эпидермы и слоя паренхимных клеток. Осевой цилиндр чайного корня состоит из перицикла, паренхимных клеток и пучка проводящей системы, образующей ксилему и флоэму. В зависимости от характера почвы чайных плантаций по-разному располагается и корневая система. На глубоких красноземах у полновозрастных насаждений основная масса корней находится в горизонтах до 50—70 cм, на подзолах — 40—60 см.
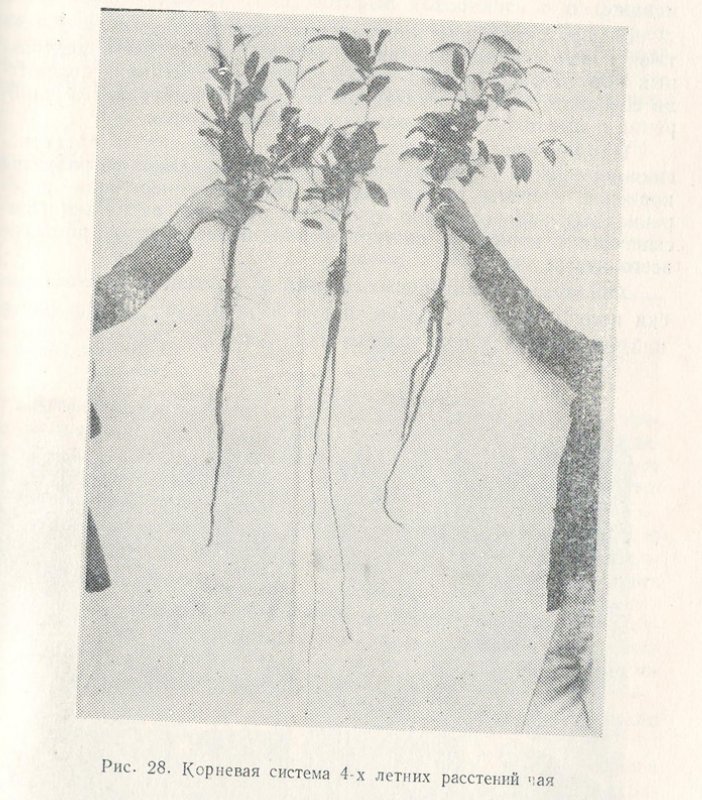
По всей длине стержневого корня находятся боковые проводящие корни 1-го порядка длиной от 20—50 см до 1,2—1,4 м. Главная масса как активных, так и проводящих корней чайного куста на красноземах расположена неравномерно. Наибольшая концентрация активных корней (70—75%) находится в почвенных слоях до 30—50 см; в более глубоких слоях почвы размещается не более 20—23% корней. При проведенных нами раскопках было установлено, что некоторая часть корней на красноземах проникает в почву на глубину до 2 м и ниже.
В горизонтальном направлении корневая система захватывает всю площадь питания, однако больше всего деятельных корней расположено в междурядьях плантаций, где всегда сосредоточены в достаточном количестве минеральные удобрения и имеется лучший водно-воздушный режим.
Применение различных видов минеральных и органических удобрений оказывает существенное влияние на развитие корневой системы чайного куста. Полное минеральное удобрение вызывает особенно усиленное развитие активных (всасывающих) корней и, соответственно, более мощное развитие всего куста.
Общая масса активных корней у растений, систематически получающих удобрения, в два раза больше, чем у растений, недостаточно получающих их.

Корневая система чайного куста характеризуется цикличностью своего развития. У активных корней в годовом цикле отмечены три периода роста и два периода покоя. Весенний период роста захватывает конец февраля, март и апрель месяцы. В это время отмечается массовое отрастание новых всасывающих корней и одновременно продолжается интенсивный рост корней прошлого года, которые находились в состоянии зимнего покоя. Начало жизнедеятельности корней совпадает с температурой почвы в верхних горизонтах 7°—9°С. В надземной части растения этому периоду соответствует набухание, одиночное, а затем массовое распускание листовых почек.

Второй период роста корней летний — июль-август, а третий — осенний, продолжающийся с сентября до половины декабря. В надземной части куста этим периодам соответствует усиленный рост побегов 1—2—3—4-х порядков, а также бутонизация, цветение и завязывание семян.
Периоды покоя приходятся на конец мая-июнь месяцы и на конец декабря, январь и начало февраля, когда рост корней не отмечен даже при самых благоприятных условиях среды (оптимальная температура, влажность почвы и воздуха). Так, если в апреле рост корней достигал 60—66% по отношению к росту в годовом цикле, то в мае он уже колебался в пределах 15—25%, в июне имеет место общая приостановка роста корней.
Продолжительность летнего периода покоя корней обычно бывает не более 30—40 дней.
Зимний покой в субтропиках побережья в отличие от всех предыдущих периодов характеризуется почти полным прекращением вегетации надземной и корневой части растения.
Соответственно с периодами роста и покоя должна осуществляться и вся технология по уходу за чайными растениями: внесение удобрений, летняя подкормка, осенне-зимняя и летняя обработки почвы. Изучение механизма поступления воды и питательных растворов в растение имеет не только теоретический, но и практический интерес. В годовом цикле жизнедеятельности чайного куста отмечается фаза частичного отмирания активных корней, которая особенно четко прослеживается в зимний период. Подобное отмирание всасывающих корней проф. П. Г. Шитт (1940) назвал корнепадом, проводя аналогию с ежегодным листопадом. Явление корнепада отмечали также в своих работах В. А. Колесников (1962), И. А. Муромцев (1940) и другие исследователи. Отмирание корней имеет место и у других теплолюбивых вечнозеленых растений— камелий, благородного лавра, фундука и других субтропических культур. Наши исследования показали, что у взрослых растений корнепад происходит интенсивнее, чем у молодых. При неблагоприятных условиях вегетации (длительное отсутствие осадков, похолодание и промерзание почвы) корнепад на чайных кустах усиливается. Отмирание корней происходит на всех глубинах почвы приблизительно одинаково (В. В. Воронцов, 1955).
Ранней весной происходит усиленный рост новых корней, число которых значительно перекрывает массу корней, погибших во время корнепада. В месте отделения отслужившего жизнь корня образуется тонкая пробковая ткань, предохраняющая его живую часть от механических повреждений и проникновения инфекции.
Одновременно с биологическим отмиранием корневой системы в годовом цикле происходит и ее частичное уничтожение при проведении различных агротехнических приемов (вспашка, культивация, перекопка). Опыты, проведенные Ш. С. Гигиберия (1939), А. Д. Каличава (1950), С. X. Пирцхалайшвили (1947) показали, что при ежегодной зимней перекопке почвы на чайных плантациях уничтожается значительная масса как деятельных, так и проводящих корней, повреждаются корни и при летних культивациях.
По А. Д. Каличава (1950) повышение урожайности чайных плантаций, оставленных без зимней перекопки, составило в среднем за 6 лет 8%. По нашим данным на ежегодно перекапываемой плантации на глубине до 10 см имелось 2,4% всасывающих корней; в то же время в почве, необрабатываемой в том же горизонте, их находилось 8,1% от общей массы всех корней. Поэтому в технологии по возделыванию чая предусматривается рекомендация, позволяющая избежать частых обработок почвы.
Корневые волоски и микориза чайного куста
Вопрос усвоения питательных растворов и воды корнями чая имеет различные толкования. Некоторые исследователи считают, что этот процесс происходит с помощью лишь корневых волосков. Академик П. М. Жуковский (1940), С. А. Келли (1938) полагают, что чайный куст является микотрофным растением.
В. В. Воронцовым установлено, что на поверхности всасывающих корней чайного куста очень мало корневых волосков, а иногда они совсем отсутствуют. В среднем на 1 см всасывающего корня их приходится до 89 штук, в то время как у яблони на этой же площади их имеется около двух-трех тысяч.

Длина имеющихся корневых волосков чая крайне незначительна и колеблется в пределах нескольких десятков микрон, в зоне корневого чехлика их нет вообще, как это имеет место у других растений. В то же время на кончиках корня чайных растений просматривается большое количество гиф, число которых достигает 15—20 на 1 мм корня. Грибница однородна, имеет небольшое количество перегородок, цвет гиф варьирует от светло-желтого до темно-желтого оттенков.
Гифы гриба распространяются по межклетникам и проникают в отдельные клетки коркового слоя; хорошо прослеживаются везикулы шарообразной формы диаметром от 20 до 40 микрон. Развитие грибницы в тканях корня, а также на нежных их окончаниях позволило нам установить наличие у чайного куста эктоэндотрофной микоризы, находящейся в симбиозе с растением-хозяином и выполняющей роль в снабжении растений водой и питательными элементами.
Отношение чайного растения к условиям произрастания
Для получения высоких и устойчивых урожаев кусты чая требуют определенных условий внешней среды, отвечающих их природе. Несоответствие этих элементов приводит к угнетению или гибели растений или вызывает необходимость разработки специальных агротехнических мероприятий, направленных на изменение условий жизнедеятельности.
Сумма активных температур, при которых происходит нормальная вегетация чайного растения, составляет 3000°—4000°С.
М. Ф. Харебава (1965), сравнивая термические ресурсы советских субтропиков с Китаем, Японией, Францией, пришел к выводу, что в наших субтропических районах температура в период вегетации ниже, суммы активных температур меньше, а зимы значительно суровее, чем в названных странах. Среднегодовая температура воздуха на Черноморском побережье Западной Грузии находится в пределах 12,5°—14,7°С, а в период вегетации —16,5°—23,9°С. Температурным барьером для начала и окончания вегетации является +10°С.
Растения чая особенно требовательны к теплу в период вегетации. Наиболее интенсивное отрастание побегов происходит при температуре 20°—25°С. Длина вегетационного периода у него составляет примерно 220—230 дней. Сумма активных температур в районах возделывания промышленных насаждений чая должна быть не менее 3000°—4000°С.
В зимний период фаза покоя в советских субтропиках у чайных растений ясно выражена даже при высоких зимних температурах. Ростовой покой обычно начинается с окончанием вегетативного роста и понижением среднесуточных температур до 8°—10°С. На севере субтропиков полное прекращение вегетации происходит в конце ноября.
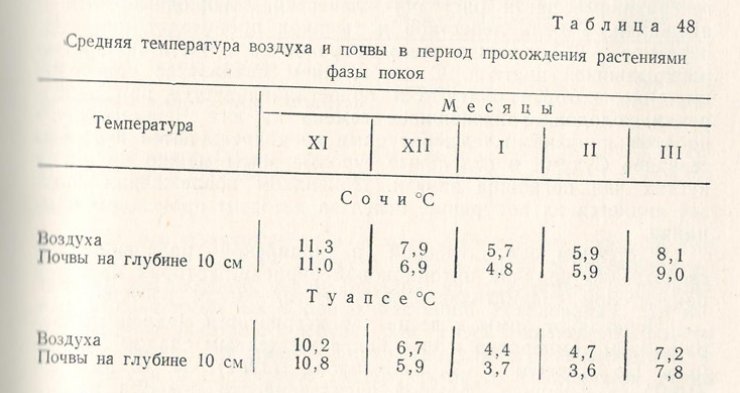
Однако полной приостановки всех жизненных процессов у растения чая в это время не происходит, они продолжают ассимилировать, дышать, бутонизировать и цвести.
Ранне-осенние заморозки даже сравнительно небольшой силы способны вызвать гибель значительной части нежных листьев и молодых неодревесневших побегов. Причиной повреждений чайных кустов могут быть сильные снегопады, сопровождаемые поломами скелетных ветвей и сучьев. Глубокий снежный покров, особенно в предгорьях Сочи—Лазаревское, приводит к выпреванию листьев и их гибели. Условием успешной перезимовки чая является прохождение в осенне-зимний период соответствующей закалки, которая позволяет растениям в фазе ростового покоя сравнительно лето переносить понижения температуры.
В значительной степени благополучное состояние растений зимой обусловлено своевременным вызреванием древесины, достаточным количеством запасных пластических веществ, в первую очередь, сахаров и жиров, концентрацией клеточного сока, а также рельефом местности, высотой над уровнем моря, типами почвы, оптимальными сроками проведения агротехнических мероприятий. Наиболее морозостойкие растения чая китайского вида в условиях Западной Грузии легко переносят низкие температуры даже без снежного покрова. Незначительные повреждения иногда у них отмечаются в виде гибели нежных растущих побегов и побурении листьев. При —6°, —8°С повреждаются листья и иногда части однолетнего прироста. Гибель растений до корневой шейки происходит при —15°, —20°С. Индийская группа растений, как правило, вымерзает при снижении температуры до —6°, —8°С.
На понижение температуры в первую очередь реагируют генеративные части растений — лепестки, тычиночные нити и пыльники. Гибель лепестков и тычинок происходит при температуре —2°С, прежде всего повреждаются части полностью раскрывшихся цветков. С потеплением начинается повторное цветение, которое опять-таки приостанавливается при повторении холодов. Завязавшиеся семена на юге Черноморского побережья низкими температурами не повреждаются и только севернее Сухуми в отдельные суровые зимы можно видеть на кустах чая погибшие завязи. Признаком повреждения завязей является их побурение, вслед за которым происходит осыпание.
Растениям чая китайской и индийской разновидностей свойственно наличие антоциановой окраски, которая является показателем сравнительной морозоустойчивости растений.
Переход от зимы к весне характеризуется медленным нарастанием температур и еще более медленным спадом их осенью. По данным Г. Т. Селянинова (1961), А. С. Мосияш (1961), в условиях большой расчлененности рельефа микроклиматические вариации отмечаются не только в температуре воздуха, но и в температуре почвы.
Требования чая к теплу в периоды бутонизации и цветения, в сравнении с другими субтропическими культурами, относительно невысокие. Если лимон цветет в мае при среднемесячной температуре 17°—19°С, то интенсивное цветение чая отмечается в октябре-январе при среднемесячной температуре 6°—9°С.
Закладка цветочных почек у чая происходит в конце июня— начале июля, когда температура воздуха поднимается до 18°—20°С. В этот период растениям особенно необходимо достаточное питание и водоснабжение.
По мере продвижения чайных плантаций на север у растений под воздействием низких температур и изменяющихся почвенных условий отмечаются сокращение срока вегетации, уменьшение размеров кроны и снижение урожайности зеленого листа. Из приведенных данных видно, что уже в Адлере средние размеры кроны куста на 17 см меньше, чем в Чакве. Еще севернее, в Лазаревском совхозе, эта разница по ширине кроны достигает более 40 см. В самом северном Гойтхском отделении Дагомысского совхоза крона чайного куста в два раза меньше, чем в Чакве.
Отношение к свету. Несмотря на то, что чай является растением подлеска в условиях Черноморского побережья, он не боится прямой солнечной радиации и не нуждается в затенении, как это практикуется в жарких тропических странах, где для получения высоких урожаев чайного листа применяют притенение насаждений путем посадки на плантациях специальных высокоствольных бобовых растений. Установлено, что ночью чайное растение требует для своей жизнедеятельности меньше света, чем днем. Однако при слабой освещенности, в особенности в пасмурную погоду, реакция фотосинтеза резко ослабевает и поступление пластических веществ в растение заметно снижается.

Требования чая к почвенным условиям. Сравнительно широкий опыт по возделыванию чайных насаждений Западной Грузии, Азербайджана, Черноморского побережья Краснодарского края показывает, что высокий урожай листа можно получать лишь на глубоких, хорошо аэрируемых почвах, которые должны отвечать следующим основным требованиям:
- быть глубокими, кислыми (pH водной суспензии 4,5—5,5) до глубины 70—80 см; ·не иметь избытка влаги;
- обладать хорошей структурой и высокой водо- и воздухопроницаемостью, которые в значительной степени зависят от механического состава почвы.
Лучшими для чая являются красноземы и бурые лесные почвы, богатые перегноем и питательными веществами.
Желтоземные и желтоземно-подзолистые почвы имеют кислую реакцию, бедны гумусом и питательными веществами, часто переувлажнены, в нижней части профиля иногда оглеены. На отдельных участках встречаются ортштейновые горизонты, и такие почвы требуют проведения больших мелиоративных работ, обогащения органическим веществом, глубокого рыхления с применением киллиферования. Чаще всего встречаются они на равнинах Колхиды или на некоторых склонах приморской полосы Абхазской АССР, Зугдидекого района, а также на верхних надпойменных аккумулятивных террасах.
Большое количество желтоземных почв имеется в Ленкоранской зоне Азербайджанской ССР. В значительной своей части указанные почвы характеризуются бесструктурностью и малой мощностью пахотного горизонта. На глубине 15—20 см на этих почвах встречаются зерна ортштейна и ортштейновые слои мощностью до 20—40 см, а ниже появляются оглеенные горизонты с ярко выраженным сизо-серым оттенком. В отдельных хозяйствах горизонты ортштейна имеют толщину 0,5—0,7 м.
В результате слабой просачиваемости нижних горизонтов выпадающие осадки в осенне-зимнее время приводят к поверхностному застою воды (верховодка), которая в период сильных дождей сплошь и рядом смыкается с близко подходящими грунтовыми водами, образуя в зоне деятельности корневой системы длительное заболачивание. Продолжительное нахождение корневой системы чая в воде без доступа кислорода вызывает загнивание и гибель большой ее массы. В результате ежегодной гибели части корней от вымокания происходит постепенное перемещение основной массы корневой системы из нижних почвенных горизонтов в верхние. В этом случае замедляется рост кустов, резко снижается урожайность.
Чайные растения, подвергающиеся длительному воздействию воды, имеют поверхностную корневую систему, которая в засушливые периоды сезона не обеспечивает их водой и минеральным питанием.

В особенно сильные засухи на таких почвах полностью прекращается вегетация, а в условиях Краснодарского края и Азербайджанской ССР кусты сбрасывают листья; в отдельные годы происходит повреждение скелетных побегов, как это имело место в Краснодарском крае в засуху 1957 и 1986 годов.
Чайные растения, заложенные на желтоземных почвах, без длительного окультуривания и проведения мелиоративных мероприятий даже при сравнительно благоприятных условиях вегетационного периода приносят низкий урожай. Так, по данным В. В. Какауридзе и др. (1985), общая масса деятельных корней полновозрастных чайных растений на желтоземах в основном расположена в слое почвы 0—40 см.
Не останавливаясь на общеизвестных приемах борьбы с избыточным увлажнением как для чая, так и для других культур, укажем на самые эффективные способы:
- глубокое рыхление междурядий не менее, чем на 40—45 см с обязательным разрушением плотно слежавшихся слоен почвы и взламыванием ортштейна. Эта работа должна выполняться плантажными плугами или киллифером — рыхлителем на тяге мощных тракторов.
- внесение органических удобрений (торфа, компоста, органических остатков сельскохозяйственного производства), рыхление почвы способствует созданию благоприятных ее водно-физических свойств. Органические удобрения вносятся при рыхлении с дальнейшей их заделкой на возможно большую глубину.
- самым главным моментом при освоении тяжелых почв и процессе их окультуривания является устройство собирательных коллекторов, сброс с участков избыточной воды и 2-х—3-х летнее окультуривание путем посева однолетних злаковых и бобовых трав и ежегодная заделка их на глубину 20—30 см.
Источник: М.К. Дараселия, В.В. Воронцов, В.П. Гвасалия, В.П. Цанава. Культура чая в СССР. «Мецниереба». Тбилиси. 1989












