Немного о происхождении китайских и индийских чайных растений на основе данных генетического анализа.
Несмотря на большое внимание к чаю, некоторые вопросы ботаники чайных растений остаются неясными либо неоднозначными. Можно констатировать наличие генотипически и фенотипически различающихся разновидностей, которые разные учёные в разное время относили или к разным вариантам единого биологического вида, или к разным биологическим видам. Раньше выделяли крупнолистную прямоствольную Camelia assamica, распространённую на юго-западе Юньнани (а также на северо-востоке Индии), мелколистный кустарник Camelia sinensis, преобладающий в других чаеводческих регионах Китая, и промежуточную камбоджийскую форму — Camelia assamica subsp. lasiocalyx. Сейчас более популярен другой подход – рассматривать культивируемые человеком прямоствольные чайные камелии как Camelia sinensis var. assamica, а мелколистные – как Camelia sinensis var. sinensis. Но эти разновидности существуют не только в чистом виде – многие сорта чайных растений, как найденные в природе и размноженные, так и выведенные искусственно, являются гибридами, содержащими генетический материал и var. assamica, и var. sinensis. Кроме того, в предгорьях Гималаев существует большое число дикорастущих камелий, и некоторые из них – например, Camelia taliensis – определённо имеют отношение к происхождению ныне культивируемых чайных растений и даже сами иногда используются для производства чая.
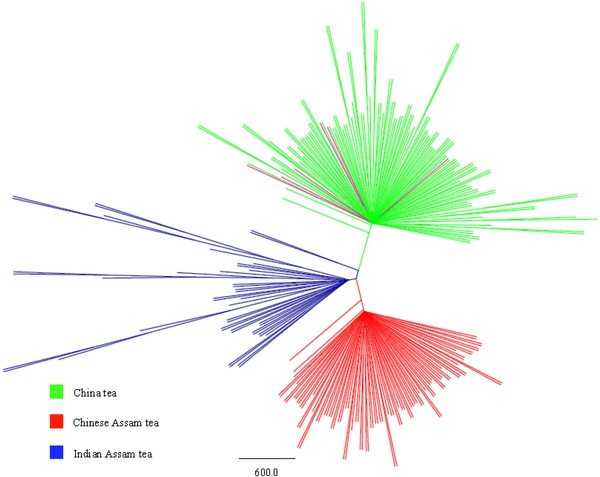
Рисунок 1. Разделение образцов на три группы по результатам анализа микросателлитов (розовое внутри зелёного — синенсис из Индии).
Вопрос о происхождении ассамики и синенсиса, его сроках и механизмах долгое время оставался открытым. Бытовало представление о том, что ассамика – это природная разновидность, найденная на юго-западе Юньнани и распространившаяся затем вдоль реки Янцзы, а синенсис — разновидность, искусственно выведенная более 1000 лет назад путём селекции, направленной в сторону удобства сбора, более высокой урожайности и более тонкого вкуса и аромата. В пользу такой версии говорит отсутствие дикорастущих популяций синенсиса. Но убедительных доказательств у этой гипотезы нет.
В 2010-х годах большая группа китайских, индийских, цейлонских, кенийских и британских учёных под руководством М.K. Meegahakumbura провела ряд исследований, которым посвящены публикации http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0155369 и https://www.frontiersin.org/articles/10.3389/fpls.2017.02270/full. Было изучено свыше 400 образцов чая из Индии и 14 основных чаеводческих провинций Китая, относящихся к природным и искусственно выведенным сортам Camelia sinensis var. sinensis, Camelia sinensis var. assamica и Camelia taliensis. Проводилось генотипирование микросателлитов ядерной ДНК (микросателлиты – короткие участки ДНК, состоящие из тандемно повторяющихся мономеров, они не кодируют белки, не подвергаются действию естественного отбора и обладают высокой изменчивостью. Поэтому микросателлиты широко используются в генетике для изучения эволюционных процессов, определения принадлежности образца к той или иной конкретной популяции и т.п.) с использованием набора из 23 праймеров, а также изучение трёх областей ДНК хлоропластов. Затем полученная информация обрабатывалась несколькими разными методами статистического анализа.
Результаты свидетельствуют о наличии в истории как минимум трёх независимых событий одомашнивания чая. Синенсис, китайская ассамика и индийская ассамика – три разные генетические группы с высокой степенью дифференциации микросателлитов между группами (рисунок 1). Шесть образцов синенсиса из Индии генетически близки к китайским и отдельной группы не образуют — по-видимому, эти сорта сформировались в Китае и были заимствованы оттуда. Камбоджийская (по морфологическим признакам) разновидность также не образует отдельную группу и демонстрирует смешанный состав генетического материала, в котором признаки индийской ассамики преобладают над признаками китайской ассамики – очевидно, камбоджийская разновидность возникла в результате скрещивания этих двух типов, произошедшего сравнительно поздно.
Во всех группах был отмечен высокий уровень генетического разнообразия. Особенно высок он был у индийской ассамики, что, вероятно, связано с более интенсивной искусственной гибридизацией, и, возможно, значительная доля генетического материала холодостойких китайских разновидностей сыграла важную роль в успешной адаптации чая к высокогорью Дарджилинга. В целом, почти треть всех образцов обладала смешанным генотипом – скрещивание разных линий было важнейшим селекционным подходом. Среди гибридных образцов встречались как гибриды F1 и F2 (чаще всего), так и беккросс-гибриды (продукты скрещивания гибрида с одним из родителей). Таким образом, в морфологическом синенсисе часто присутствует генетический материал ассамики (причём не только китайской – иногда и индийской), и наоборот; во многих образцах китайской ассамики обнаруживается генетический материал Camelia taliensis; в ДНК хлоропластов некоторых образцов индийской ассамики есть последовательности, соответствующие диким видам Camelia pubicosta.
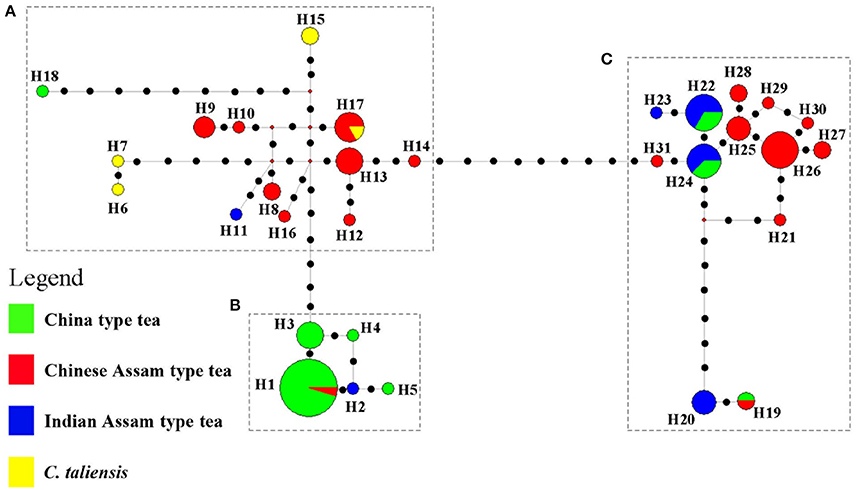
Рисунок 2. Разделение образцов по результатам анализа ДНК хлоропластов.
Демографическое моделирование, в котором продолжительность одного поколения была принята равной 12 лет, показало, что синенсис и ассамика впервые разошлись порядка 22500 лет (1875 поколений) назад, что примерно совпадает с последним ледниковым периодом. По-видимому, в результате изменения климата область распространения будущих чайных камелий сместилась к югу, и это послужило толчком для их эволюции. Вмешательство человека в этот процесс маловероятно. А вот момент расхождения индийской ассамики и китайской ассамики – примерно 2770 лет (231 поколение) назад – близок к началу употребления чая в Китае.
Анализ ДНК хлоропластов китайской ассамики позволяет разделить её на две подгруппы. Ассамика с юга Юньнани (Сишуаньбаньна и Пуэр (бывший Сымао)) группируется вместе с Camelia taliensis (на рисунке 2 слева), а ассамика с западной границы Юньнани (Линьцан и Баошань) близка к индийской ассамике (на рисунке 2 справа). Это заставляет авторов предположить, что западноюньнаньская ассамика имела другого материнского родителя (хлоропласты передаются по материнской линии), нежели южноюньнаньская, а общность их микросателлитов объясняется интенсивным потоком генетического материала между субрегионами Юньнани, что размывает первоначальные генетические границы.
Точно локализовать одомашнивание синенсиса на данный момент невозможно из-за отстутствия диких популяций.
Следует отметить также, что эти исследования лишний раз подтвердили, что типирование чайных растений по внешним признакам не всегда даёт правильный результат, и в ряде случаев результаты генетического анализа позволяют исправить ошибки – 7 из 392 образцов в первой работе первоначально были идентифицированы неверно. Морфологические особенности чайных камелий изменчивы и зависимы от факторов окружающей среды – говоря простым языком, внешний вид чайных растений, относящихся к одной и той же разновидности, может отличаться настолько, что по нему не всегда можно точно определить, какая разновидность перед вами.
10 июля 2018 г.
Источник: Самая домашняя чайная «Сова и Панда» https://vk.com/club47905050
Автор: Антон Дмитращук https://vk.com/id183549038












